Типы прикрепления хромосом к кинетохорам
Обновлено: 15.05.2024
Веретено деления — динамичная структура, которая образуется в митозе и мейозе для обеспечения сегрегации хромосом и деления клетки. Типичное веретено является биполярным — между двумя полюсами образуется веретенообразная система микротрубочек. Микротрубочки веретена присоединяются к кинетохорам хроматид в области центромер и обеспечивают движение хромосом по направлению к полюсам.
Веретено образуют три основных структурных элемента:
- микротрубочки,
- полюса деления,
- хромосомы.
В организации полюсов деления у животных участвуют центросомы, содержащие центриоли. У растений, а также в ооцитах некоторых животных центросомы отсутствуют, и образуется ацентросомальное веретено с широкими полюсами. Важную роль в формировании веретена играют моторные белки, относящиеся к семействам динеинов и кинезинов.
Полноценное веретено деления образуется на стадии прометафазы после разрушения ядерной мембраны, когда цитоплазматические микротрубочки и центросомы (у животных) получают доступ к хромосомам и другим компонентам веретена. Исключение составляет веретено деления почкующихся дрожжей, которое формируется внутри ядра.
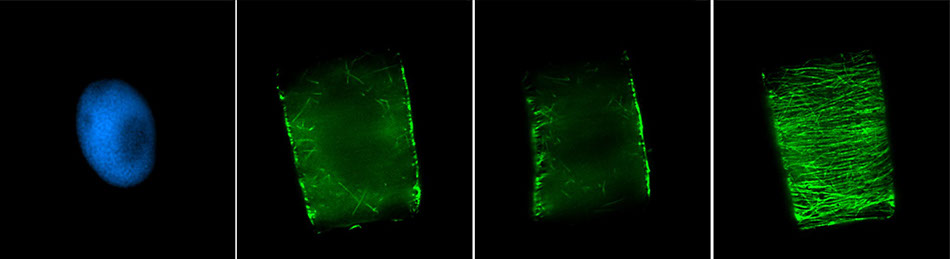
Веретено деления типичной клетки млекопитающих состоит из трёх структурных элементов — центросом, микротрубочек и хромосом, — которые образуют симметричную биполярную структуру. На полюсах веретена располагаются центросомы — небольшие органеллы, функционирующие как центры организации микротрубочек. Каждая центросома состоит из пары центриолей, окруженных множеством разных белков. Между полюсами веретена находятся конденсированные хромосомы, состоящие из пары хроматид, скреплённых в области центромеры. На цетромерных участках хромосом находятся кинетохоры — сложные структуры, отвечающие за прикрепление к микротрубочкам веретена.
Веретено деления состоит из двух полуверетён. Полуверетено образуется из поляризованных микротрубочек. Отрицательные минус-концы микротрубочек собираются на полюсах веретена вокруг центросом. Плюс-концы микротрубочек отдаляются от двух полюсов и пересекаются в средней экваториальной части веретена. У большинства позвоночных полуверетено состоит из 600—750 микротрубочек, 30—40 % которых заканчиваются на кинетохорах. Микротрубочки, которые соединяют полюса веретена с кинетохорами хромосом, называются кинетохорными. Причём каждый кинетохор при образовании веретена связывается с множеством микротрубочек и образует кинетохорный пучок. Микротрубочки, которые располагаются между полюсами и не присоединяются к кинетохорам, называются межполюсными. Часть микротрубочек веретена образует вокруг каждого полюса радиальные структуры, называемые звёздами или астерами. Такие микротрубочки называются астральными.
У растений, а также в ооцитах некоторых животных центросомы отсутствуют, и образуется ацентросомальное веретено с широкими полюсами. Также на полюсах ацентросомального веретена отсутствуют астральные микротрубочки. В остальном структура веретена растительной клетки соответствует структуре веретена животной клетки.
Сборка веретена деления начинается в профазе. Однако на данном этапе образование полноценного веретена невозможно по причине изоляции хромосом, а также важных моторных, регуляторных и стабилизирующих белков внутри ядра.
У растений, по причине отсутствия центросом, роль центра организации микротрубочек в профазе выполняет ядерная оболочка. Микротрубочки собираются вблизи поверхности ядра и к окончанию профазы ориентируются вдоль оси будущего веретена деления, образуя так называемое профазное веретено.
В животных клетках центром организации микротрубочек является центросома. Поэтому образование веретена деления начинается с разделения и расхождения пары центросом во время профазы. Расхождение центросом в профазе обеспечивают моторные белки динеины. Они закрепляются на внутренней стороне клеточной мембраны и на внешней поверхности ядра. Закреплённые в мембране динеины присоединяются к астральным микротрубочкам и движутся в направлении минус-конца микротрубочки. За счёт этого центросомы перемещаются к противоположным участкам клеточной мембраны и расходятся дальше друг от друга.
У всех эукариот сборка биполярного веретена деления зависит по большей части от способности компонентов веретена к самоорганизации. Самоорганизация — единственный механизм сборки веретена деления в клетках лишённых центросом. Сборка биполярного веретена без участия центросом называется ацентросомальной. Она характерна для высших растений, а также наблюдается при мейозе на ранних стадиях развития некоторых животных. Более того, предполагается, что самоорганизация микротрубочек является преобладающим механизмом сборки веретена, даже в животных клетках, содержащих центросомы.
- Нуклеация микротрубочек вблизи хромосом.
- Соединение и сортировка антипараллельных микротрубочек при участии кинезина-5.
- Кинезины-4 и -10, закреплённые на плечах хромосом, также участвуют в сортировке и отдаляют минус-концы микротрубочек.
- Минус-концы микротрубочек скрепляются между собой и образуют полюса деления при участии динеина и кинезина-14.
Сборка веретена зависит от двух ключевых процессов. Во-первых, от формирования биполярного скопления микротрубочек вокруг хромосом. Во-вторых, от прикрепления хромосом к микротрубочкам от противоположных полюсов деления. Присоединение сестринских хроматид к микротрубочкам является неотъемлемой частью процесса сборки веретена. Однако, хромосомы и многие моторные и другие белки, участвующие в формировании полноценного веретена деления изолированы внутри клеточного ядра. А микротрубочки и центросомы (у животных) находятся в цитоплазме. Таким образом, сборка веретена зависит от разрушения ядерной оболочки в прометафазе.
Во многих животных клетках, включая человеческие, в сборке веретена участвуют центросомы, являющиеся полюсами веретена деления. Также, как и при сборке ацентросомального веретена, моторные и другие белки участвуют в самоорганизации микротрубочек в биполярную структуру, которая фокусируется с помощью минус-концов микротрубочек в области центросом. Центросомы при этом тоже участвуют в сборке веретена и способствуют формированию полюсов деления, но не являются неотъемлемым компонентом веретена, так как процесс сборки может протекать даже при инактивации центросом.
преподаватель - профессор, д.б.н Е.А. Смирнова
История открытия митоза. Митоз как стадия клеточного цикла. Биологический смысл митоза. Стадии митоза - профаза, прометафаза, метафаза, анафаза, телофаза и цитокинез. Основные события в профазе митоза. Преобразование структуры хромосом в ходе подготовки к митотическому делению. Конденсины, когезины, шугошины. Механизмы, обеспечивающие когезию хромосом в митозе. Инициация сборки митотического веретена. Биполярное митотическое веретено, его структура, состав и организация. Астральное и анастральное веретено, сходства и отличия. Функции мультимерных моторных белковых комплексов в организации митотического веретена в профазе. Когда и почему может формироваться монополярное веретено Прометафаза, основные события ранней и поздней прометафазы. Разрушение ядерной оболочки в прометафазе, судьба белков ламинов и ядерной мембраны. Центромерный район хромосомы и кинетохор. Структура и молекулярный состав кинетохора. Варианты прикрепления кинетохоров к микротрубочкам веретена. Активное и пассивное состояние кинетохоров. Исправление ошибочных вариантов прикрепления. Комплекс СРС, его молекулярный состав, локализация, участие в контроле прикреплении микротрубочек к кинтохору. Механизмы ассоциации микротрубочек и кинетохоров - поиск и захват (search and capture), кооперативное взаимодействие между микротрубочками, отходящими от полюсов и от кинетохоров. Генерирование силы в области кинетохора. Вклад хромосом и кинетохоров в создание структуры митотического веретена. Роль градиента RanGTP в нуклеацию микротрубочек от хромосом и кинетохоров, участие в этом процессе белков NuMa, TPX2 и моторных белков. Вклад аугминовых комплексов в формировании митотического веретена. Формирование митотического веретена без участия центросомы. Вклад само-организации микротрубочек в формирование веретена. Анастральное веретено. Силы, направляющие движение хромосом к полюсам и экватору веретена во время прометафазы. Конгрессия хромосом и укладывание в метафазную пластинку. Осциллирование хромосом в прометафазе-анафазе. Перетекание тубулина (flux) и его функции в митозе. Пункты контроля правильности прикрепления хромосом к микротрубочкам веретена. Инициация разделения хроматид в начале анафазы. Анафаза А и B. Механизм движения хромосом в анафазе А. Роль кинетохора в движении хромосом. Вклад в этот процесс моторных белков и деполимеризации микротрубочек. Расхождение полюсов в анафазе В и механизмы этого процесса. Основные события во время телофазы. Цитокинез и его стадии. Генерирование сигнала от центросомы и микротрубочек в клеточный кортекс. Роль митотического веретена в определении плоскости деления. Белки центросомы и кинетохоров и их роль в регуляции формирования контрактильного кольца. Централспиндлиновый комплекс и СРС, их участие в цитокинезе. Структура и организация контрактильного кольца. Механизм его сокращения. Мембранный трафик во время цитокинеза. Механизм разделения дочерних клеток. Патологии митоза. Классификация патологических митозов, характеристика К-митозов, многополюсных и монополярных митозов. Причины возникновения разных форм патологии митоза. Физические и химические способы индукции патологических митозов.
Особенности митотического деления клеток высших растений. Сходства и отличия цитоскелета растительных и животных клеток. Микротрубочки. Биохимические особенности микротрубочек клеток высших растений. Тубулин растительных клеток, изоформы растительного тубулина и пост-трансляционные модификации тубулина. Белки ассоциированные с микротрубочками (MAP), особенности растительных белков семейства МАР. Белки семейства +TIP, их роль в регуляции динамики микротрубочек и ассоциации микротрубочек с различными органеллами. Моторные белки растений: кинезины, миозины, из молекулярные и функциональные особенности. ЦОМТ-независимая нуклеация микротрубочек растительных клеток. Роль γ-TURC в нуклеации микротрубочек. Ассоциация γ-TURC с латеральной поверхностью микротрубочек. Особенности динамики микротрубочек растений: гибридный тредмиллинг. Организация системы микротрубочек высших растений в клеточном цикле. Интерфазная кортикальная и радиальная сеть. Структура, организация и функции. Возможные механизмы регуляции упорядоченной организации кортикальной сети, реорганизация кортикальной сети в ответ на внешние воздействия. Роль кортикальных микротрубочек в отложении фибрилл целлюлозы при формировании клеточной стенки. Связь между кортикальными микротрубочками и движением целлюлоза-синтазных комплексов. Фрагмосома вакуолизированных клеток. Эндоплазматические/радиальные микротрубочки, их роль в заякоривании ядра и его пред-митотической миграции. Препрофазное кольцо (ППК) как часть цитокинетического аппарата клеток растений: с чем связано присутствие ППК в определенных типах дифференцированных клеток. Корреляция между расположением ППК, осью митотического веретена и плоскостью деления клеток. ППК в вакуолизированных и невакуолизированных клетках. Структурная и функциональная связь между эндоплазматическими/радиальными микротрубочками и ППК. Стадии формирования ППК. Пространственная связь между ППК, веретеном и фрагмопластом. Происхождение микротрубочек, формирующих ППК. Модели формирования ППК. Взаимоотношения между ППК, фрагмосомой и фрагмопластом. Роль актиновых филаментов в определения положения ППК и его организации. Формирование доменов на плазматической мембране в зоне расположения ППК. Модели организации кортикальной зоны деления. Функции ППК. Митотическое веретено высших растений как типичный пример анастрального веретена. Стадии формирования веретена в клетках с ППК и без ППК. Пластичность веретена растительных клеток, разные типы организации анастрального веретена у растений. Кинетохорная фибрилла как структурная основа веретена растений. Типы микротрубочек в составе кинетохорной фибриллы. Белковый состав веретена. γ-TURC растений и участие γ-тубулина в нуклеации микротрубочек веретена. Пластичность полюсов веретена, их трансформация во время митоза. Установление биполярности веретена в отсутствие полюсных ЦОМТ. Стадии дифференцировки веретена во время его формирования в профазе-прометафазе. ЦОМТ-независимый механизм формирования митотического веретена растений, роль само-организации микротрубочек и моторных белков. Участие в сборке веретена градиента RanGTP и моторных белков. Цитокинетический аппарат растительный клеток. Отличия цитокинетического аппарата растительных и животных клеток. Стадии цитокинеза у растений. Фрагмопласт, его структура, расположение и роль в формировании клеточной пластинки между дочерними ядрами. Латеральный рост фрагмопласта. Происхождение микротрубочек фрагмопласта в разных типах растительных клеток (имеющих ППК и без ППК). Динамика микротрубочек в составе фрагмопласта. Механизмы формирования фрагмопласта и клеточной пластинки. Белки, обнаруженный в составе фрагмопласта, их роль в формировании, поддержании структурной целостности, обеспечении динамических преобразований и функциях фрагмопласта. Везикулярный транспорт во время цитокинеза. Стадии созревания клеточной пластинки. Актин и актин-связывающие белки растений. Нуклеация актиновых филаментов у растений. Актиновые филаменты в клеточном цикле разных типов клеток. Субкортикальные пучки в интерфазе, их роль в циклозе. Динамика актиновых филаментов в растительных клетках. Роль актина в определении плоскости деления клеток (Actin Depletion Zone). Расположение актиновых филаментов во время митоза и цитокинеза. Актин в составе веретена и фрагмопласта, его участие в поддержании структуры фрагмопласта и везикулярном транспорте. Роль актиновых филаментов в процессах внутриклеточного транспорта растений. Опосредованное миозином перемещение стопок аппарата Гольджи по актиновым филаментам вдоль цистерн ЭПР. Координация процессов внутриклеточного транспорта, осуществляемого при участии микротрубочек и актиновых филаментов.
• Часто в процессе прикрепления хромосомы к веретену происходят случайные ошибки, которые исправляются
• Ошибочные прикрепления неустойчивы, поскольку они не позволяют кинетохорам стабилизировать прикрепившиеся микротрубочки
• Только правильное биполярное прикрепление хромосомы приводит к стабильному прикреплению кинетохора
Чувствительность места контакта микротрубочки и кинетохора к натяжению играет важную роль в коррекции ошибок прикрепления, которые происходят при формировании веретена. Механизм поиска-захвата может служить источником двух типов ошибок прикрепления, которые могут происходить в ходе процесса нормального митоза.
Хромосома прикрепляется синтелически (от греч. syn — тот же самый; telos — конец), когда оба ее кинетохора прикрепляются к одному и тому же полюсу. Ошибка обычно происходит сразу после разрушения оболочки ядра, когда хромосомы находятся в случайной ориентации и могут оказаться гораздо ближе к одной центросоме, чем к другой. Это дает возможность сестринским кинетохорам одновременно захватывать микротрубочки, исходящие из одной и той же центросомы.
Один кинетохор может прикрепляться сразу к двум полюсам (меротелическое прикрепление; греч. mero — часть). Хромосомы, содержащие один (или даже два) меротелически ориентированных кинетохора, нормально движутся к экватору веретена и могут представлять серьезную опасность для клетки. Если меротелическое прикрепление сохраняется, две хроматиды разделяются при вступлении клетки в анафазу, но ошибочно прикрепившаяся хроматида остается у экватора веретена.
Она находится там до тех пор, пока не нарушится ее связь с одним из полюсов, что обеспечит 50%-ю вероятность ее сегрегации к тому же полюсу, что и сестринская хроматида.

Биориентация является единственным типом прикрепления, который обеспечивает одинаковое распределение хромосом по двум дочерним клеткам;
все хромосомы должны быть биориентированы до начала анафазы.
Моноориентация представлет собой нормальный промежуточный этап на пути к биориентации.
В ранней прометафазе часто наблюдается синтелическое и меротелическое прикрепление,
однако до наступления метафазы эти типы превращаются в биориентированное прикрепление.
Обычно синтелические и меротелические прикрепления исправляются вскоре после того, как они образовались. Для обоих типов ошибочных прикреплений характерно, что нить кинетохора расположена не под прямым, а под острым углом к кинетохору. Это приводит к повреждению структуры кинетохора и к дестабилизации его связи с пучком микротрубочек.
В результате микротрубочки начинают отделяться быстрее, а замещаться медленнее, чем при правильном прикреплении. Это приводит к тому, что в конце концов в нити кинетохора больше не остается микротрубочек, и связь разрушается. В зависимости от типа ошибочного прикрепления, утрата связи приводит или к исправлению ошибки за счет создания биориентированной хромосомы, или к появлению хромосомы, прикрепленной только к одному полюсу через один кинетохор. Такая моноориентированной хромосома может затем по обычному механизму превратиться в биориентированную.
Важную роль в исправлении ошибочных прикреплений также играет чувствительность кинетохоров к натяжению. Ошибки прикрепления не позволяют кинетохорам обращаться к полюсу, тем самым предотвращая развитие натяжения, необходимого для их правильного прикрепления. Особенно характерна нестабильность для синтелического прикрепления, поскольку в этом случае на два кинетохора приходится незначительное натяжение, существенно меньшее, чем при правильной ориентации хромосомы и локализации центромеры между двумя полюсами.
Такая ошибка часто исправляется или при спонтанной потере одной из связей из-за отсутствия натяжения, или когда один из двух кинетохоров дополнительно прикрепляется к отдаленному полюсу. При этом в направлении полюса развивается резкое усилие, которое нарушает структуру кинетохора и дестабилизирует его прикрепление.
Чрезвычайная запутанность механизма прикрепления демонстрирует всю структурно-функциональную сложность кинетохоров и их центральное место в процессе митоза. Фактически расхождение реплицированных хромосом на две одинаковые группы непосредственно связано с функцией кинетохоров. В 1961 г. Мэзия — один из пионеров исследования механизмов митоза — обратил внимание на их исключительную роль. Он писал, что кинетохор «представляет собой единственную существенную часть митотической хромосомы».
Мэзия сравнивал остальную часть хромосомы с «трупом на похоронах» поскольку она «служит причиной события, но не принимает в нем непосредственного участия». Он был прав, считая, что истинная цель митоза состоит в сегрегации сестринских кинетохоров (остальную часть хромосомы можно рассматривать лишь как ездовые аксессуары). Однако в то время он не мог предвидеть, насколько глубоко вовлечены в этот процесс кинетохоры.
Итак, нити кинетохора прикрепляют хромосому к веретену и определяют направление ее движения. Одинаковая сегрегация хромосом в митозе может быть непосредственно связана со следующими установленными фактами:
(1) каждая реплицированная хромосома содержит два сестринских кинетохора;
(2) эти кинетохоры расположены на противоположных сторонах первичной перетяжки и обращены в противоположных направлениях;
(3) кинетохорные микротрубочки связываются только с его внешней частью, направленной в сторону от хромосомы: и
(4) существуют механизмы исправления ошибок прикрепления кинетохоров к полюсам. Отсутствие этих механизмов приводит к анеуплоидии.

Возможность исправления неправильного прикрепления хромосомы
зависит от динамических свойств микротрубочек в митозе и от чувствительности кинетохоров к возникновению у них различных нарушений.
Только при достижении конфигурации, изображенной внизу, может сформироваться нормальная нить кинетохора, и хромосома надежно связывается с веретеном.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Читайте также: